

Physiology of Pruning Fruit Trees
ID
422-025 (SPES-235P)
 EXPERT REVIEWED
EXPERT REVIEWED
Introduction
Woody plants are pruned to maintain a desired size and shape and to promote a certain type of growth. Ornamental plants are pruned to improve the aesthetic quality of the plant, but fruit trees are pruned to improve fruit quality by encouraging an appropriate balance between vegetative (wood) and reproductive (fruiting) growth. Annual pruning of fruit trees always reduces yield, but enhances fruit quality. Pruning increases fruit size because excess flower buds are removed and pruning encourages the growth of new shoots with high-quality flower buds. Pruning improves light penetration into the canopy, and light is required for flower-bud development, fruit set and growth, and red color development. Pruning also makes the canopy more open and improves pest control by allowing better spray penetration into the tree; air movement throughout the canopy is increased, which improves drying conditions and reduces severity of many diseases.
This publication describes why plants respond to pruning and other forms of plant manipulation used to train trees. This information applies to all plants, but application to fruit trees is emphasized.
Pruning fruit trees is somewhat of an art based on an understanding of plant physiology and development. In other words, if we understand how plants grow and how they will respond to different types of plant manipulations, we can alter vegetative growth and fruiting to obtain trees and fruit with desirable characteristics.
A basic understanding of certain aspects of plant physiology is a prerequisite to understanding pruning. Unlike animals, plants continue to increase in size throughout their lives. There are only two ways plants can grow.
Primary growth is the increase in length of shoots and roots, and is responsible for increases in canopy height and width.
Secondary growth is the increase in thickness of stems and roots.
Both types of growth require cell division followed by cell enlargement and differentiation.
Plant Growth
Meristems are regions of cell division and there are two types of plant meristems. An apical meristem is located at the tip of every shoot and root (Figure 1). As cells divide in these apical meristems, the shoots and roots elongate as cells are piled one on another. Behind the region of cell division is a region of cell differentiation, where cells enlarge and differentiate into various tissues. In the axil of each leaf is a small apical meristem called an axillary meristem that forms an axillary bud, which usually remains dormant until well after the subtending leaf is fully developed. An axillary bud may remain dormant or develop into a lateral branch or a flower.
There are two distinct layers of meristematic tissue within the stem or root responsible for secondary growth, the vascular cambium and the cork cambium (Figure 2). The vascular cambium is a cylinder of specialized cells, usually five to ten cells thick, running the length of the plant, including the roots, and is responsible for the radial growth of plant parts. Phloem cells are produced to the outside of the cambium and xylem cells are produced to the inside of the cambium.

Downward transport of sugars, nutrients, and hormones from the top of the tree to the roots occurs in the phloem tissue. Xylem cells are tube shaped, become hollow and die to form a pipe-like system through which water, hormones and mineral nutrients move from the roots to the top of the tree. Most of the radial growth of woody plants is due to activity of the vascular cambium, but a small amount results from activity in another lateral meristem, the cork cambium, located outside the vascular cambium. The cork cambium (phellogen) together with the cork cells, constitute the periderm: a protective layer of suberized dead cork cells forming the bark. Suberization is the impregnation of cell walls of cork tissue with a fatty substance called suberin. Each season new layers of cells are produced and appear as growth rings when viewed in cross-section. Over time, the xylem cells at the center of the trunk or limb are crushed and become nonfunctional as transport pipes, but they do provide structural support to hold the plant upright. While grafting it is important to line up the cambiums of the scion and the rootstock to ensure a successful graft union.
Buds
Buds are important to the vegetative and reproductive growth of trees. Fruit tree training and, to a lesser extent, pruning primarily involves bud manipulation. Buds are actually undeveloped shoots. When a vegetative bud is sliced longitudinally during the winter and viewed under magnification, the apical meristem at the tip, leaf primordial (developing leaves), axillary meristems, developing axillary buds, and procambial tissue (tissue that will develop into the cambium) are all visible.

Buds on fruit trees usually have about seven leaves and initial shoot elongation in the spring results from cell expansion. During late June and July some of the shoot apices will flatten out and develop into flower buds. Flower buds are actually modified shoots and the various flower tis- sues (petals, stigmas, anthers, etc.) are actually modified leaves. Although the process of switching from vegetative to reproductive buds is not fully understood, hormones that can be influenced by environmental factors, stresses, and plant nutrition control the process.
There are several things we can do to influence whether or not a bud becomes a flower bud or remains vegetative. In general, factors that favor rapid growth, such as high nitrogen levels in the shoot tissues, inhibit the development of flower buds. Applying growth-promoting plant growth regulators such as gibberellins usually inhibits flower-bud induction, whereas ethylene may promote flower-bud development. Mild stresses such as shoot bending and water stress may also promote flower-bud development.
Producing annual crops of high-quality fruit requires a balance between reproductive and vegetative growth. Fruit producers use various tech- niques, including pruning, branch bending, and plant growth-regulator sprays, to manipulate tree growth and flowering. Often these techniques affect bud dormancy, so knowledge of buds and bud dormancy is essential if we are to understand how pruning influences tree growth. It is also important to be able to identify the different types of buds on a tree, especially to distinguish between flower and vegetative buds.
Buds may be classified as to location, contents, or activity.
Classification by content
Several types of buds commonly develop on fruit trees. Vegetative buds only develop into leafy vegetative shoots. Flower buds produce only flowers. Stone fruit trees (peach, nectarine, apricot, plum, and cherry) produce vegetative buds and flower buds. Apple and pear trees produce vegetative and mixed buds. Both leafy shoots and flowers emerge from mixed buds.
Classification by location
Terminal buds are located at the tip of a shoot. On stone fruit trees terminal buds are vegetative buds. Terminal buds on apple and pear trees are usually vegetative; however, some varieties such as ‘Rome Beauty’ produce mixed buds terminally and are referred to as “tip bearers” or “terminal bearers.” Most mixed buds on apple and pear trees are formed terminally on short, less than six-inch, shoots that terminate with a rosette of leaves. These short shoots are called “spurs.” Lateral buds form in the axils of leaves and are often referred to as “lateral buds” or “axillary buds.” On stone fruit trees, lateral buds may be either vegetative or flower. Nodes on one-year-old shoots may have one to three buds, some of which may be flower buds and others vegetative buds. Flower buds are larger with tips that are relatively round, whereas vegetative buds are small, narrow, and pointed. In the case of apple and pear trees, lateral buds on the previous season’s growth are usually vegetative. However, lateral buds on some varieties, especially on the dwarfing rootstocks, may be mixed buds.
Classification by arrangement on the stem
The bud arrangement influences the arrangement of a fruit tree’s branches and thus the tree’s shape and how easy it is to manage. A node is the joint on a stem where a leaf is or was attached (Figure 3). Axillary buds are located in the axis above where a leaf is attached to the stem. In apples there is usually only one leaf per node, whereas three leaves often arise from a node on peach shoots. When a leaf falls in the autumn, a leaf scar remains just below the axillary bud (Figure 3). Buds are opposite when there are two at the same node but on opposite sides of the stem. Forsythia is an example of a plant with opposite buds. Buds are alternate when there is only one from each node and no one bud is on the same side of the stem as the one next above or below it. Deciduous fruit trees have buds that spiral along a shoot (Figure 4). The spiraling three-dimensional arrangement of leaves around a stem is known as Phyllotaxy and is expressed as a fraction, where the numerator is the number of turns to get to a leaf directly above another and the denominator is the number of buds passed.


Classification by activity
Buds are dormant when they are not visibly growing. When shoots develop around large pruning cuts, they usually are sprouting from dormant buds (Figure 5). Adventitious buds form irregularly on older portions of a plant and not at the stem tips or in the leaf axils. They form on parts of the root or stem that have no connection to the apical meristems. They may originate from either deep or peripheral tissues. For example, shoots often arise from adventitious buds growing from callus tissue around wounds. Root suckers (vigorous upright shoots developing from the roots) develop from adventitious buds on the roots.

Plant Hormones
Hormones are substances produced in very small amounts in one part of the plant and transported to another part where they cause a response. Plants produce a number of hormones that control various aspects of growth, such as stem elongation; dormancy of buds and seeds; flowering; fruit set, growth, and ripening; and the response to light and gravity. While pruning, it is useful to consider the activity of the general types of hormones, “promoters” (gibberellins and cytokinins) and “inhibitors” (auxins and abscisic acid). Promoters generally cause bud growth, cell division and elongation, and stem growth. Inhibitors are usually associated with dormancy and inhibit shoot development from seeds and buds and may be involved in flower- bud induction. It is often the ratio of promoters and inhibitors, rather than their absolute concentrations, that determines how a plant will grow. The production of plant hormones is usually controlled by environmental conditions such as temperature or day length. Vegetative growth is usually associated with low ratios of inhibitors to promoters and dormancy is usually associated with high ratios of inhibitors to promoters.
Dormancy is a condition characterized by temporary growth cessation and suppressed metabolism. During the winter trees appear not to be growing, but the tissues are alive, there is metabolic activity, and cells are slowly expanding and differentiating. By early October all the flower parts (petals, stigmas, anthers, etc.) can be seen in a flower bud and vegetative buds contain leaves. During the winter these various tissues continue to enlarge and differentiate. Given favorable growth conditions, some buds will develop into shoots or flowers, but others may remain dormant. By understanding the factors influencing bud dormancy we often can influence certain aspects of tree growth. Buds of deciduous trees go through several stages of dormancy.
Results from dormancy research are confusing because plant physiologists have used different terminology to describe the stages of dormancy.
Plant physiologists currently describe dormancy in four stages.
Para-dormancy occurs in the mid to late summer when buds do not grow because inhibitors produced in the leaves and terminal buds inhibit bud growth. Para-dormancy can often be overcome by removing leaves (leaf stripping) along a section of a shoot so the axillary buds develop into shoots. Nurserymen often use this technique to produce trees with lateral branches (feathered trees). Using heading cuts to remove the terminal portion of a shoot will allow several axillary buds just below the cut to develop into shoots. Sometimes an application of growth promoters (gibberellins and/or cytokinins) will induce bud growth.
Sometimes axillary buds do not become dormant and develop into shoots within a few days of being formed. Such shoots are referred to as sylleptic shoots and are fairly common on vigorously growing peach trees, but are rarely produced on apple trees (Figure 6).
Ecto-dormancy occurs in the early fall, before defoliation, when plants do not grow because the environmental conditions are not conducive for growth. Growth will resume if the plants are exposed to suitable temperatures and day lengths.
Endo-dormancy occurs during the winter because there are high levels of inhibitors (abscisic acid) within the buds. During this phase of dormancy the trees will not grow even under ideal growing conditions. The concentration of inhibitors declines as buds are exposed to chilling temperatures. Temperatures near 45˚F are ideal for chilling, but temperatures between 35˚ and 55˚ F will provide some chilling. The chilling requirement to satisfy dormancy for most varieties of apples and peaches grown in Virginia is about 1,000 and 800 hours, respectively. When the chilling requirement is satisfied, the level of inhibitors within the bud is low enough that growth may commence when environmental conditions are appropriate for growth. Avoid planting varieties with chilling requirements less than 800 hours because such varieties usually bloom early and are susceptible to frost.
Eco-dormancy occurs in the late winter, usually by mid January, after the chilling requirement has been satisfied. At this time the trees do not grow because conditions are unsuitable for growth. Growth will commence when trees are exposed to warm temperatures.
Apical dominance is a type of para-dormancy, where axillary bud growth is inhibited in the api cal meristematic zone. Axillary buds on fruit trees typically remain dormant for a prolonged period while the main shoot continues to grow. Apical dominance has been studied for more than 80 years, and the exact mechanism is not yet fully understood, but it seems to be controlled by the relative concentrations of inhibitors and promoters. Growth of axillary buds is inhibited by high concentrations of auxin produced by the terminal bud. Auxin moves down the shoot, from cell to cell by gravity, so concentrations are highest near the shoot tip. Promoters are produced in the roots and are transported upward in the tree. Growth of axillary buds may occur at the base of shoots where concentrations of inhibitors are relatively low and concentrations of promoters are relatively high.

You can overcome apical dominance by removing the shoot tip, which is the source of auxin (Fig- ure 7). The three or four buds immediately below a heading cut usually develop into shoots. Pinching annual plants to induce branching is a form of heading. Another way to overcome apical dominance is to notch buds. Notching involves cutting through the bark to hard wood, with a knife or hacksaw blade at about bloom time, just above a bud. The cut interrupts the downward flow of inhibitors, but not the upward flow of promoters, and releases the bud from dormancy. On vigorous upright one-year-old shoots, notching often successfully over-comes dormancy in about 70 percent of the buds. Sometimes apical dominance can be overcome by spraying shoots with promoters (gibberellins and/ or cytokinins) just before bloom time.

Shoot bending
Shoots bend in response to an auxin gradient within the shoot. Everyone who has grown plants in the house has noticed that plants tend to grow towards the light. This phenomenon is known as photomorphism and is caused by varying concentrations of auxin in different sides of a stem or shoot. Auxin causes cells to elongate, but auxin is destroyed by light. Therefore, there is a higher concentration of auxin on the dark side of a shoot and the cells on the dark side elongate more than cells on the sunny side of the shoot, causing the shoot to bend towards the light (Figure 8).
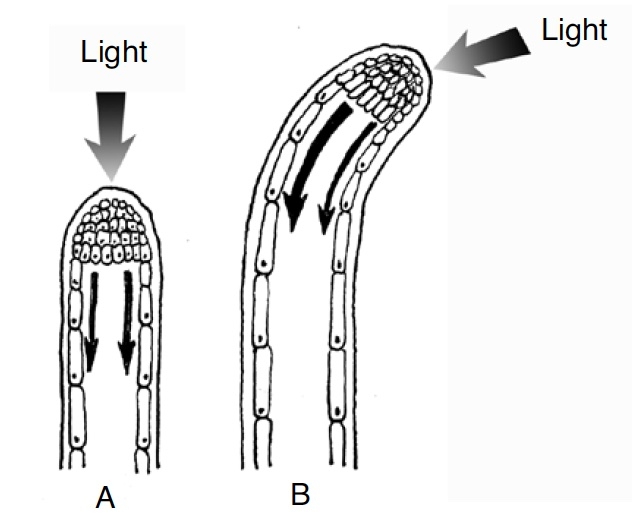
Tree fruit producers have noticed a similar phenomenon where the tips of growing branches tend to bend upward, even when the branch was physically oriented to the horizontal. This condition, known as gravimorphism, is also caused by an auxin gradient within the branch in response to gravity. Auxin flows by gravity to the lower side of a limb. The subsequent accumulation of auxin is responsible for increased cell elongation on the underside of the limb, and the growing tip bends upward.
Another consequence of gravimorphism is the development of watersprouts from the upper surface of horizontally oriented limbs. Watersprouts are vertically growing shoots that develop from the upper surfaces of branches or near pruning cuts. High auxin concentrations on the underside of the limb inhibits growth of buds on the underside of the limb, but the concentration of auxin on the upper side of the limb is inadequate to inhibit bud growth and many of these buds develop into watersprouts. Watersprouts are usually undesirable and their development can be suppressed by orienting limbs no more than 45 degrees from the vertical. Fruit trees are sometimes trained as espalier (tree fence). There are several ways to espalier trees, but one method involves orienting limbs to a horizontal position. This system induces many watersprouts along the length of the branches. Watersprout development can be greatly suppressed by orienting limbs 45 to 60 degrees above horizontal (Figure 9).
Reducing tree height by cutting into large diameter branches or trunks often results in the development of vigorous watersprouts around the cut. There are buds buried in the bark that normally remain dormant. However, a severe pruning cut will release these buds from dormancy.

Additional Pruning Facts
Pruning is a dwarfing process
Pruning increases vegetative growth near the pruning cut and this gives the illusion that pruning stimulates growth. However, the weight of a tree that was pruned annually is always less than the weight of a nonpruned tree.
Pruning reduces yield
Pruning removes wood with flower buds, and thus potential fruit. Yield from pruned trees is nearly always less than yield from nonpruned trees, but fruit quality is improved by pruning. Pruning improves fruit size by increasing the amount of leaf area per fruit. Pruning improves light distribution throughout the tree, which is important for the development of fruit red color and sugar levels.
Pruning delays fruiting
Pruning encourages vegetative growth rather than reproductive growth in young trees. A nonpruned tree will always flower and produce fruit earlier in the life of the tree than a pruned tree. The reason young trees are pruned is to induce branches to develop where they are wanted and to develop a strong tree structure that will support large crops as the tree matures. As a tree matures the physiology changes from vegetative growth to reproductive growth. To obtain high annual yields of mature trees, it is important to minimize fruiting until trees have nearly filled their space. Pruning is one technique used to delay fruiting of young trees.
Summer pruning
Summer pruning involves the selective removal of leafy shoots during the growing season. Responses to summer pruning vary with time of pruning, severity of pruning, tree vigor, geographical location, and variety. Several researchers evaluated summer pruning during the 1980s and several general statements can be made about the practice.
Summer pruning reduces within-tree shade and usually improves fruit red color development and sometimes improves flower bud development.
Summer pruning removes leaves that produce pho- tosynthates (sugars) for growth of all tree parts. Summer pruning sometimes reduces fruit size and sugar levels.
Due to reduced whole-tree photosynthesis, summer pruning suppresses late-season trunk enlargement and root growth.
Summer pruning does not suppress shoot elongation the following season. Summer pruning reduces late-season photosynthesis, and theoretically should reduce the accumulation of reserve carbohydrates within the tree that are used for early season growth. However, results from most pruning experiments indicate that the response to a certain type of pruning cut will be the same regardless of the time of year the cut was made.
Summary
Pruning is an important orchard practice because pruning can influence fruit quality and the balance between vegetative growth and fruiting. Successful pruners observe how plants respond to various types of plant manipulation, including pruning. Profitable fruit production requires an understanding of plant physiology, and how pruning alters the physiology of the plant. For further information concerning how to prune fruit trees, see the Extension publications that provide information on how to prune and train apple and peach trees.
Training and Pruning Apples Trees, Virginia Cooperative Extension publication 422-021, http://pubs.ext.vt.edu/422-021/
Training and Pruning Apple Trees in Intensive Orchards, Virginia Cooperative Extension publication 422-024, http://pubs.ext.vt.edu/422-024/
Pruning Peach Trees, Virginia Cooperative Extension publication 422-020, http://pubs.ext.vt.edu/422-020/
Virginia Cooperative Extension materials are available for public use, reprint, or citation without further permission, provided the use includes credit to the author and to Virginia Cooperative Extension, Virginia Tech, and Virginia State University.
Virginia Cooperative Extension is a partnership of Virginia Tech, Virginia State University, the U.S. Department of Agriculture (USDA), and local governments, and is an equal opportunity employer. For the full non-discrimination statement, please visit ext.vt.edu/accessibility.
Publication Date
September 1, 2025
